
A D-allulóz élettani funkcióival és bioszintézisével kapcsolatos legújabb eredmények
Allulose StoreOldal meghallgatás
Forrás: frontiers
Élelmiszer-mikrobiológia szak
13. kötet - 2022 |https://doi.org/10.3389/fmicb.2022.881037
Ez a cikk a kutatási témája 'A részeInnovatív mikrobiális technológiák a jövőbeli és fenntartható élelmiszertudományért'-nek
A D-allulóz élettani funkcióival és bioszintézisével kapcsolatos legújabb eredmények
- Szénhidrátkémiai és Biotechnológiai Kulcslaboratórium, Oktatási Minisztérium, Biotechnológiai Iskola, Jiangnan Egyetem, Vuxi, Kína
A D-allulóz, egy általánosan biztonságosnak (GRAS) tartott cukor, a természetben ritka.
Alacsony kalóriatartalma, szacharózszerű íze és egyedi funkciói miatt a jövőbeni felhasználásra az egyik legígéretesebb édesítőszer.
A D-allulóznak számos fiziológiai hatása van, például elhízás elleni, hiperglikémia elleni, cukorbetegség elleni, gyulladáscsökkentő, antioxidáns és neuroprotektív hatása van.
Ezért a D-allulóz fontos alkalmazási értékkel bír az élelmiszeriparban, a gyógyszeriparban és az egészségügyben.
A D-allulóz előállításának magas költsége azonban korlátozza nagymértékű alkalmazását. Jelenleg a biotranszformáció nagyon vonzó a D-allulóz szintézise szempontjából, a bioszintézis két fő módszere az Izumoring stratégia és a DHAP-függő aldoláz stratégia.
Ez a cikk áttekintést nyújt a D-allulóz fiziológiai funkcióival és bioszintézisével kapcsolatos legújabb eredményekről. Ezenkívül bemutatja a D-allulóz előállításának jövőbeli kilátásait.
Bevezetés
Az utóbbi időben az elhízás, a hiperlipidémia, a magas vérnyomás és a cukorbetegség kockázata világszerte gyorsan megnőtt a magas zsír- és cukortartalmú, tápláló étrendek túlzott bevitele miatt.
A szacharóz, egy hagyományos élelmiszer-édesítőszer, édessége és ízletessége miatt fontos szerepet játszik az élelmiszeriparban ( Castro-Muñoz et al., 2022 ).
A szacharóznak azonban van néhány hátránya, például magas kalóriatartalma, hiperglikémiás reakciók kiváltására való képessége és a cukorbetegség ( Grassi et al., 2021 ).
Ezért az alacsony kalóriatartalmú édesítőszer-helyettesítők felkeltették a kutatók érdeklődését ( Khan et al., 2021 ).
Több mint 30 féle ritka cukrot jelentettek ( Granström et al., 2004 ). Egyedi biológiai funkciókkal rendelkeznek, és élelmiszer-adalékanyagként, ráksejt-szupresszorként, valamint rákellenes és vírusellenes gyógyszerek építőköveiként használják őket ( Zhang et al., 2016b ; Li et al., 2017 ; Guerrero-Wyss et al., 2018 ; Hoshikawa et al., 2018 ; Xia et al., 2021 ).
A D-allulóz (más néven D-pszikóz) a ritka cukrok közül a legfigyelemreméltóbb.
Több mint 20 év telt el azóta, hogy Ken Izumori először számolt be a D-allulózról ( Itoh et al., 1995 ).
Édességének 70%-át mutatja, de energiatartalmának csak 0,3%-át, mint a szacharóz.
Ezenkívül szinte semmilyen kalóriát nem tartalmaz ( Matsuo et al., 2002 ).
Ipari szempontból fontos biotermékként a D-allulózt az Egyesült Államok Élelmiszer- és Gyógyszerügyi Hivatala (FDA) 2002-ben „általánosan biztonságosnak elismert” (GRAS) anyagként sorolta fel, és jóváhagyta édességekben, gyümölcslevekben, táplálékkiegészítőkben és egyéb étrendi termékekben való felhasználását.
Ezért a D-allulóz fontos alkalmazási értékkel bír az élelmiszeriparban, a gyógyszeriparban és az egészségügyben.
A D-allulóz várhatóan megváltoztatja az édesítőszer-piacot alacsony kalóriatartalma és ízletessége miatt, mivel képes lesz versenyképes lenni más cukorhelyettesítőkkel (cukoralkoholok, például xilit, mannit és szorbitol).
A D-allulóz kereskedelmi ára várhatóan megközelíti a versengő cukorhelyettesítő édesítőszerek, például a xilit (2–5 USD/kg), a mannit (1–5 USD/kg) és a szorbitol (1–10 USD/kg) árát.
A D-allulóz a közeljövőben versenyképessé válik a nagymértékű termelésben, hasonlóan az évi több millió tonnányi fogyasztású más édesítőszerekhez.
Jelenleg a D-allulóz kibocsátását és árát korlátozó tényezők közé tartozik a kiindulási anyagok magas ára, az alacsony hozamok és az izolálás nehézsége.
Ebben a cikkben a D-allulóz élettani funkcióival és bioszintézisével kapcsolatos legújabb eredményeket foglaljuk össze és tárgyaljuk.
A D-allulóz élettani funkciói
A D-allulóz hatása a lipidanyagcserére
A D-allulózról kimutatták, hogy elhízás elleni hatással rendelkezik állatokban és emberekben a táplálékbevitel, a zsírtömeg és a zsírszövet súlyának csökkentése révén ( Kimura et al., 2017 ; Bilal et al., 2018 ).
Ezenkívül számos bizonyíték arra utal, hogy a D-allulóz növelheti az energiafogyasztást és csökkentheti a zsírlerakódást normál patkányokban ( Chung et al., 2012 ; Ochiai et al., 2014 ).
Bár a D-allulóz csökkentheti a zsírszövet súlyát, a mechanizmus továbbra sem ismert.
Csökkentheti a zsírsav-szintáz (FSA) aktivitását, és növelheti a β-oxidációt és a karnitin-palmitoiltranszferáz (CPT) aktivitását a mellékhere fehér zsírszövetében (WAT; Han et al., 2016 ).
Továbbá, az étrendi D-allulóz gátolhatja a lipogenezissel kapcsolatos acetil-CoA karboxiláz alfa (ACCα) expresszióját a mellékhere WAT-ban.
Ezenkívül az étrendi D-allulóz stimulálhatja a zsírsav-oxidációval kapcsolatos AMP-aktivált protein kináz alfa 2 (AMPKα2), a hormonérzékeny lipáz (HSL) és a peroxiszóma proliferátor aktivált receptor alfa (PPARα; Chen et al., 2019 ) expresszióját. Ezért a D-allulóz potenciális elhízásellenes tulajdonságokkal rendelkezik.
A D-allulóz antihiperglikémiás hatása
A megfelelő táplálkozás az elhízás és az elhízás okozta 2-es típusú cukorbetegség (T2DM) előfordulásának gyors növekedéséhez vezetett világszerte, ami jelentősen megnöveli ezen krónikus betegségek kezelésének költségeit.
Ezért nagyon fontos a cukorbetegség és szövődményeinek kezelésére szolgáló hatékony terápiás beavatkozások azonosítása. A D-allulóz ígéretes antihiperglikémiás tulajdonságai miatt nagy figyelmet kapott, mivel képes szabályozni a plazma glükózszintjét, a testsúlyt és a zsírtömeget ( Matsuo és Izumori, 2009 ; Hossain et al., 2015 ; Lee et al., 2020 ).
Antidiabetikus hatásának mechanizmusa azonban továbbra sem tisztázott. Feltételezések szerint a D-allulóz befolyásolja a vércukorszintet és az inzulinszekréciót, vagy a glükozidáz és a lipolitikus enzimek aktivitását.
Kimutatták, hogy a D-allulóz jelentősen gátolja a szacharóz vagy maltóz által kiváltott plazma glükózkoncentráció-növekedést ( Matsuo és Izumori, 2009 ).
Ezenkívül a D-allulózról kimutatták, hogy hatékonyan gátolja a bélszacharáz és maltáz aktivitását.
A D-allulóz (5 g) hozzáadása jelentősen elnyomhatja az orális maltodextrin (75 g) által kiváltott vércukorszint-emelkedést normál felnőtteknél.
Ezenkívül az orális D-allulóz önmagában nem befolyásolja a glükóz és az inzulin vérszintjét ( Iida et al., 2008 ).
A D-allulózról azt is kimutatták, hogy gátolja az étkezés utáni vércukorszint emelkedését, főként a határeseti cukorbetegségben szenvedő betegeknél, és 12 hét folyamatos D-allulóz-bevitelt követően nem észleltek mellékhatásokat vagy klinikai problémákat ( Hayashi et al., 2010 ).
Bár számos vizsgálat kimutatta, hogy a D-allulóz antidiabetikus aktivitással rendelkezik, a mechanizmus még nem teljesen tisztázott.
A májban a D-glükóz anyagcseréjét a glükokináz nukleocitoplazmatikus eltolódása szabályozza ( Hossain et al., 2015 ).
Alacsony glükózszint mellett a glükokináz inaktív formában marad a glükokináz szabályozó fehérjéhez (GKRP) kötődve, amelyet a hepatocita magjában toboroznak. Amikor a glükózszint eléri a magas szintet, a glükokináz a glükokináz-GKRP komplexről való leválással aktiválódik, és a magból a citoplazmába transzlokálódik, ahol részt vesz a glikogén anyagcserében és a vércukorszint homeosztázisában ( Liu et al., 2012 ).
Például a glükokináz aktiválása javíthatja a glükóz toleranciát és az inzulinérzékenységet ( Shintani et al., 2017b ).
A máj glükokináz károsodott működése a cukorbetegségben a hiperglikémia patogeneziséhez vezet ( Basu et al., 2001 ).
Ezért a glükokinázt az antidiabetikus gyógyszerek potenciális célpontjának tekintik ( Lloyd et al., 2013 ).
Ezenkívül a glükokinázt számos fruktóz-foszfát aktiválja, például a fruktóz-6-foszfát (F6P) és a fruktóz-1-foszfát (F1P; Pfefferkorn, 2013 ).
A D-allulóz-1-foszfát, hasonlóan a fruktóz-1-foszfáthoz, szintén aktiválja a glükokinázt. Ezért a D-allulóz növelheti a máj glükózhasznosítását ( Toyoda et al., 2010 ).
A glükóz-6-foszfatáz (G6Páz) hozzájárul a hiperglikémiához cukorbetegségben, és szabályozza a máj glükoneogenetikai fluxusának sebességmeghatározó lépéseit ( Herling et al., 1998 ).
A D-allulóz szabályozhatja a vércukorszintet és hipolipidémiás hatást fejt ki a G6Páz aktivitásának csökkentésével ( Nagata et al., 2015 ).
Jelentések szerint a D-allulóz orális adagolása stimulálhatja a GLP-1 szekréciót, és így alkalmazható a glükóz intolerancia megelőzésére és kezelésére ( Hayakawa et al., 2018 ).
Nemrégiben megerősítették, hogy a D-allulóz indukálja a GLP-1 felszabadulását, aktiválja a vagus afferens jelátvitelt, csökkenti a táplálékfelvételt és korlátozza a hiperglikémiát egészséges és elhízott cukorbeteg patkányokban.
Továbbá az orális D-allulóz korrigálhatja az aritmiás túlevést, az elhízást és a cukorbetegséget ( Iwasaki et al., 2018 ).
A D-allulóz egyéb élettani funkciói
A D-allulóz gyulladáscsökkentő hatást fejt ki azáltal, hogy elnyomja a gyulladást elősegítő citokinek, például a tumornekrózis-faktor-alfa (TNF-α), az interleukin-6 (IL-6) és a monocita kemoattraktáns protein 1 (MCP-1) szérumszintjét. Ezek a citokinek főként a zsigeri zsírszövetekből származnak ( Moller és Berger, 2003 ; Kim et al., 2017 ).
A D-allulóz antioxidáns hatással is rendelkezik, a reaktív oxigénfajták (ROS) megkötésével védi a 6-hidroxidopamin által kiváltott apoptózist vagy megelőzi a herekárosodást ( Takata et al., 2005 ; Suna et al., 2007 ).
A D-allulóz a szuperoxid-diszmutáz (SOD) és a kataláz (CAT) aktivitásának növelésével is meghosszabbíthatja az élettartamot ( Shintani et al., 2017a ).
Továbbá kimutatták, hogy a D-allulóz megváltoztatja a szérum koleszterinszintjét hörcsögökben, részben a proprotein konvertáz szubtilizin/kexin 9-es típusú (Pcsk9) szintjének csökkentésével ( Kanasaki et al., 2019 ).
Továbbá a D-allulóz javíthatja a szisztémás és az izom inzulinérzékenységét éber patkányokban ( Natsume et al., 2021 ). A D-allulóz különböző fiziológiai funkcióit az 1. ábra szemlélteti.
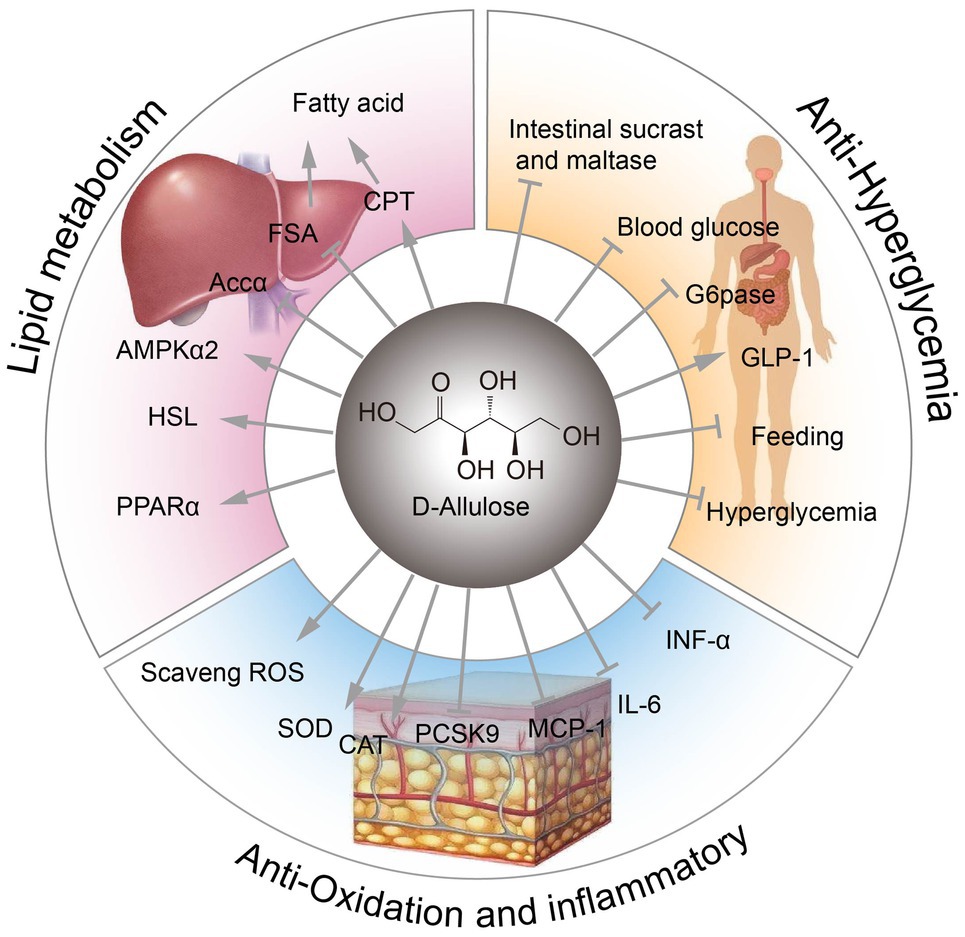
1. ábra . A D-allulóz fiziológiai funkciói. FSA, zsírsav-szintáz; CPT, karnitin-palmitoiltranszferáz; ACCα, acetil-CoA karboxiláz alfa; AMPKα, AMP-aktivált protein kináz alfa; HSL, hormonérzékeny lipáz; PPARα, peroxiszóma proliferátor aktivált receptor alfa; ROS, reaktív oxigénfajták; SOD, szuperoxid-diszmutáz; CAT, kataláz aktivitás; Pcsk9, proprotein konvertáz szubtilizin/kexin 9-es típus; MCP-1, monocita kemoattraktáns protein 1; G6Páz, glükóz-6-foszfatáz.
D-allulóz biológiai előállítása
Amint azt fentebb leírtuk, a D-allulóz számos hasznos fiziológiai funkcióval rendelkezik. A D-allulóz azonban ritka a természetben, ami nagymértékben korlátozza széleskörű alkalmazását. A D-allulóz hagyományos kémiai szintézise általában nehézkes reakciókkal és számos mellékreakcióval jár ( McDonald, 1967 ; Doner, 1979 ).
Kémiai módszerekkel nehéz egyetlen termékkonfigurációt előállítani.
Ezzel szemben a biokonverziós megközelítéseknek számos előnyük van, beleértve az enyhe reakciókörülményeket, a kevés mellékterméket, az egyszerű tisztítási lépéseket és a környezetbarát tulajdonságokat ( Zhang et al., 2021 ).
Ezért a biotranszformáció fokozatosan a D-allulóz szintézis fő módszerévé vált.
Jelenleg a D-allulóz biológiai előállítását főként két stratégiával érik el : (1) az Izumoring stratégiával és (2) a DHAP-függő aldoláz stratégiával.
D-allulóz előállítása Izumoring stratégiával
D-allulóz előállítása D-fruktózból D-tagatóz 3-epimeráz család enzimjeinek felhasználásával
Az Izumoring stratégia ígéretes megközelítés bármilyen hexózcukor biotermelésére, és D-tagatóz-3-epimerázokat (DTEázok), poliol-dehidrogenázokat és aldóz-izomerázokat foglal magában ( Izumori, 2006 ).
Az elmúlt évtizedekben az Izumoring stratégia hatékonynak bizonyult a ritka cukrok bioszintézisében.
A DTEáz családba tartozó enzimek kulcsfontosságú enzimek a D-allulóz D-fruktózból történő bioszintéziséhez ( 2. ábra ), és ide tartoznak a DTEáz és a D-allulóz-3-epimeráz (DAEáz).
A DAEáz nagyobb specificitást mutat a D-allulózra, mint a D-tagatózra ( Kim et al., 2006 ).
A Pseudomonas cichorii ST-24- ből származó első DTEáz azonosítása óta 1993-ban más DTEáz enzimeket izoláltak különféle fajokból, mint például az Agrobacterium tumefaciens ( Kim et al., 2006 ), a Clostridium Bolteae ( Jia et al., 2014 ), Dorea sp. CAG317 ( Zhang et al., 2015 ), Ruminococcus sp. 5_1_39BFAA ( Chen et al., 2016 ), Treponema primitia ZAS-1 ( Zhang et al., 2016c ), Rhodobacter sphaeroides ( Qi et al., 2017 ), Ruminococcus sp. ( Li et al., 2018 ), Arthrobacter globiformis M30 ( Yoshihara et al., 2017 ), Clostridium cellulolyticum H10 ( Su és mtsai, 2018 ), Sinorhizobium sp. ( Zhu et al., 2019b ), valamint a Rhodopirellula baltica SH 1 ( Zhang et al., 2020 ). A különböző fajokból származó DTEase-családba tartozó enzimek katalitikus tulajdonságainak részleteit az 1. táblázat mutatja be . Itoh et al. (1995) számoltak be először a DTEáz immobilizálásáról P. cichorii- ból chitopearl gyöngyökön, és 90 g D-allulózt állítottak elő 500 g D-fruktózból.
Azóta a DTEáz immobilizálási módszerei széles körű érdeklődésre tartanak számot ( Lim et al., 2009 ; Tseng et al., 2014 ; Narayan Patel et al., 2018 ), és az immobilizált enzimek jobb hőstabilitást és tárolási stabilitást mutattak
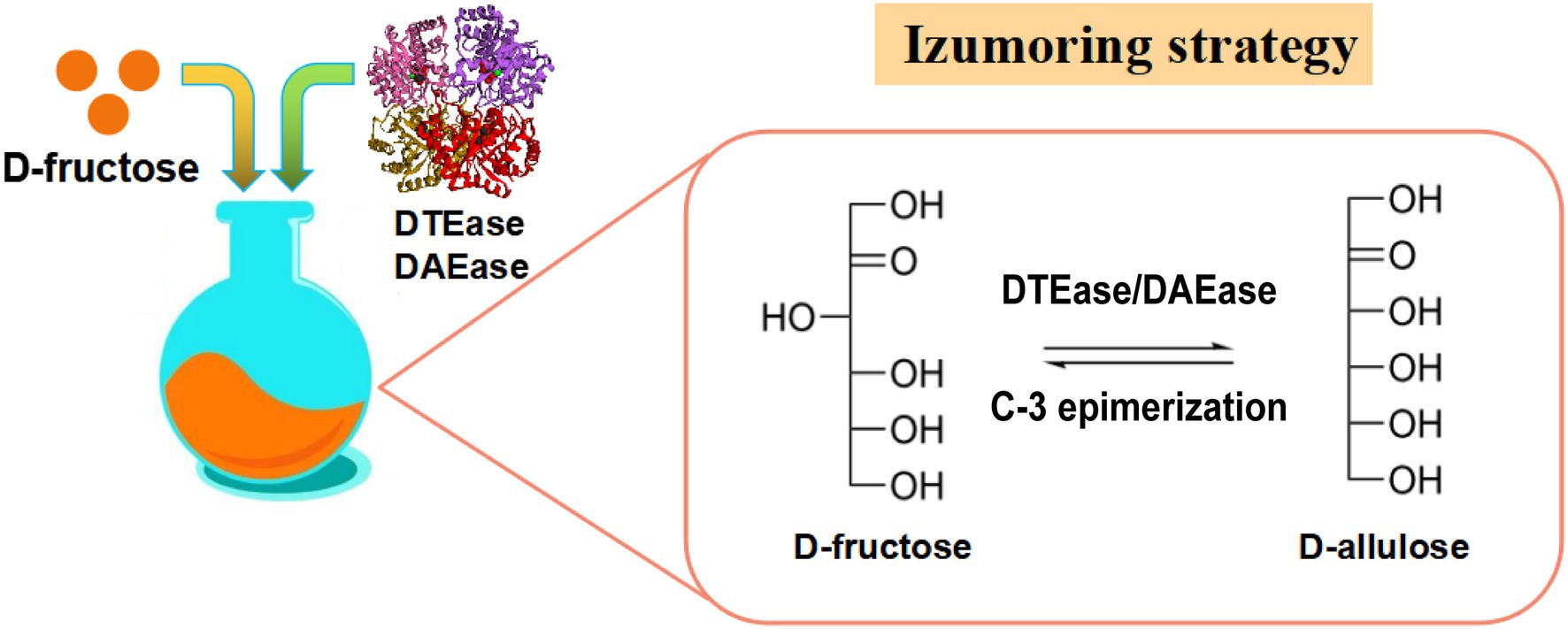
2. ábra . Izumoring stratégia D-allulóz előállítására. DTEáz, D-tagatóz-3-epimeráz; DAEáz, D-allulóz-3-epime
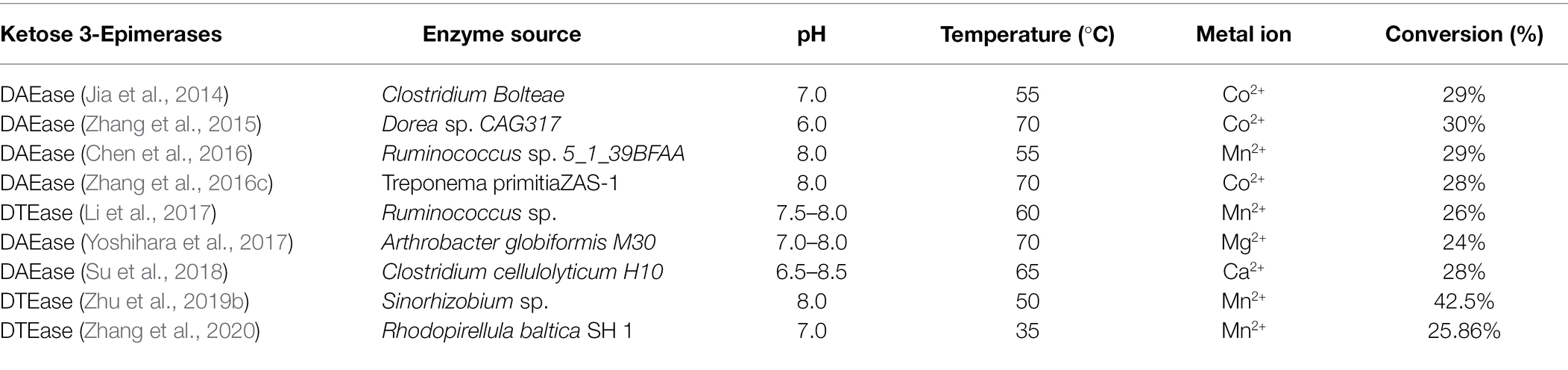
1. táblázat . A DTEáz családba tartozó enzimek tulajdonságai.
D-allulóz bioszintézise olcsó anyagokból DTEáz enzimek alapján
Jelenleg a mezőgazdasági melléktermékek, mint például a gyümölcs- és zöldségmaradványok, jelentős mezőgazdasági problémákat okoznak ( Lai et al., 2017 ).
A mezőgazdasági maradványokat általában hulladéklerakókban temetik el vagy elégetik ( Park és Yoon, 2015 ). Ezek a maradványok azonban nagy mennyiségű élelmi rostot és cukrot tartalmaznak, beleértve a szacharózt, a D-glükózt és a D-fruktózt.
Az élelmi rostok és cukrok magas hozzáadott értékű termékekké történő átalakítása jelentős lépést jelent a mezőgazdasági problémák enyhítése felé.
A kaszkádkatalízist nagyon vonzó megközelítésnek tekintik a hagyományos, lépésről lépésre történő szintézishez képest.
Ezt a stratégiát gyakran alkalmazzák ritka cukrok előállítására olcsó anyagokból, például szacharózból, csicsókából, inulinból és gyümölcs-/zöldségmaradványokból ( 3. ábra ; Wagner et al., 2015 ; Song et al., 2016 , 2017 ; Zhang et al., 2017 ; Yang et al., 2019 ; Li et al., 2020a ).
A D-allulózt hatékonyan szintetizálták szacharózból tisztított rekombináns invertáz, D-xilóz-izomeráz és DTEáz felhasználásával.
Ezenkívül a kaszkádoldat és a szimulált mozgóágyas (SMB) kromatográfia gyakorlati integrációját alkalmazták tiszta D-allulóz (99,9%) előállítására nagyon magas hozammal (89%; Wagner et al., 2015 ).
A D-allulóz kétlépéses kaszkád reakcióval is szintetizálható, amely csicsóka hidrolízist ( Song et al., 2017 ), keresztesvirágú növényi maradványokat ( Song et al., 2016 ) és inulint ( Li et al., 2020a ) foglal magában.
A termelési költségek csökkentése és az enzimtisztítás elkerülése érdekében Zhu és munkatársai egy egylépéses, két enzimes reakciórendszert fejlesztettek ki egy Bacillus velezensis (BvInu) eredetű új exo-inulinázzal és Ruminococcus sp. eredetű DAEázzal a csicsóka D-allulóz előállításához.
A BvInu-t és a DAEáz-t Bacillus subtilisben expresszálták , és tisztítás nélkül a felülúszóba szekretálták. Az optimális BvInu/RDAE arány (80:40 U/g inulin) mellett 50°C-on 2 órán át 10,3 g/l D-allulózt kaptak 50 g/l inulinból ( Zhu et al., 2020 ).
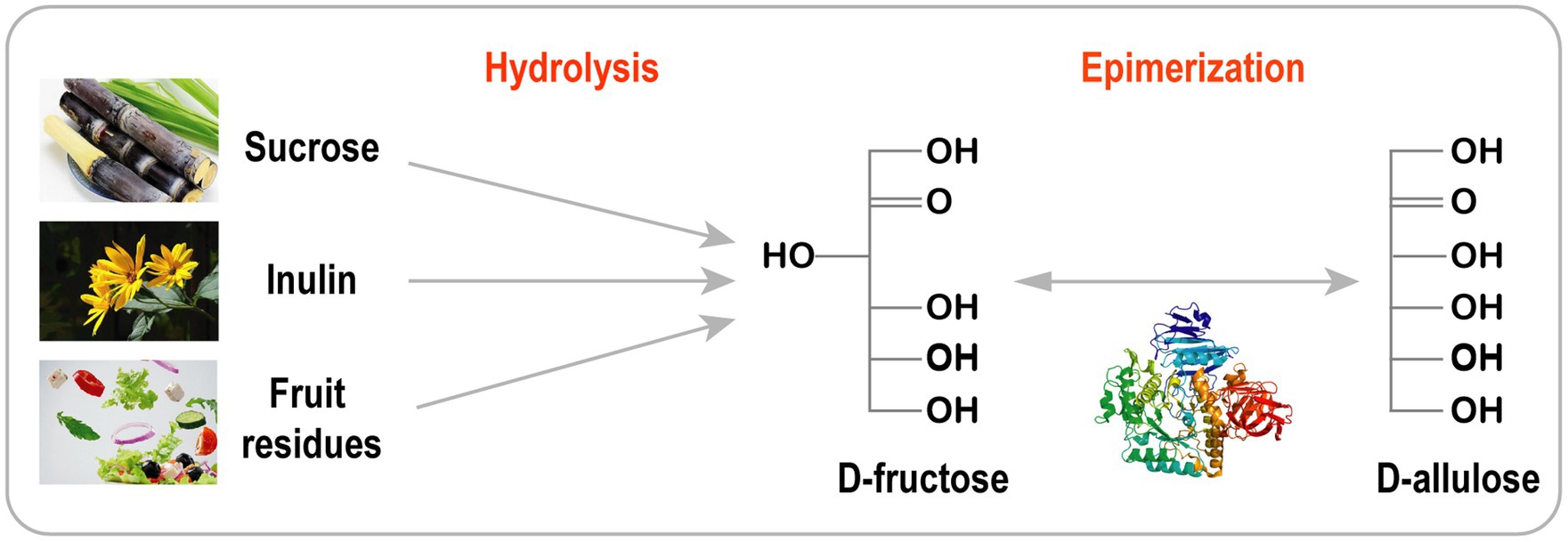
3. ábra . D-allulóz bioszintézise olcsó nyersanyagból.
D-allulóz bioszintézise mikroorganizmusok segítségével
Az egylépéses kaszkádreakciókkal összehasonlítva a teljes sejtes biokatalizátoros reakcióknak számos előnyük van ( 4. ábra ): (1) Az enzimeket tartalmazó sejtek könnyen előállíthatók az enzimek fáradságos tisztítása nélkül; (2) a sejtes kontextus megfelelő mikro-környezetet és kofaktor-regenerációt (ATP, NAD + /NADH) biztosít; (3) a sejtfalak és membránok védik az enzimeket a zord reakciókörülményektől; és (4) a több enzim sejten belüli kolokalizációja növeli az enzimek lokális koncentrációját és csökkenti az intermedierek diffúzióját a kaszkádreakciókban ( Wu és Li, 2018 ).
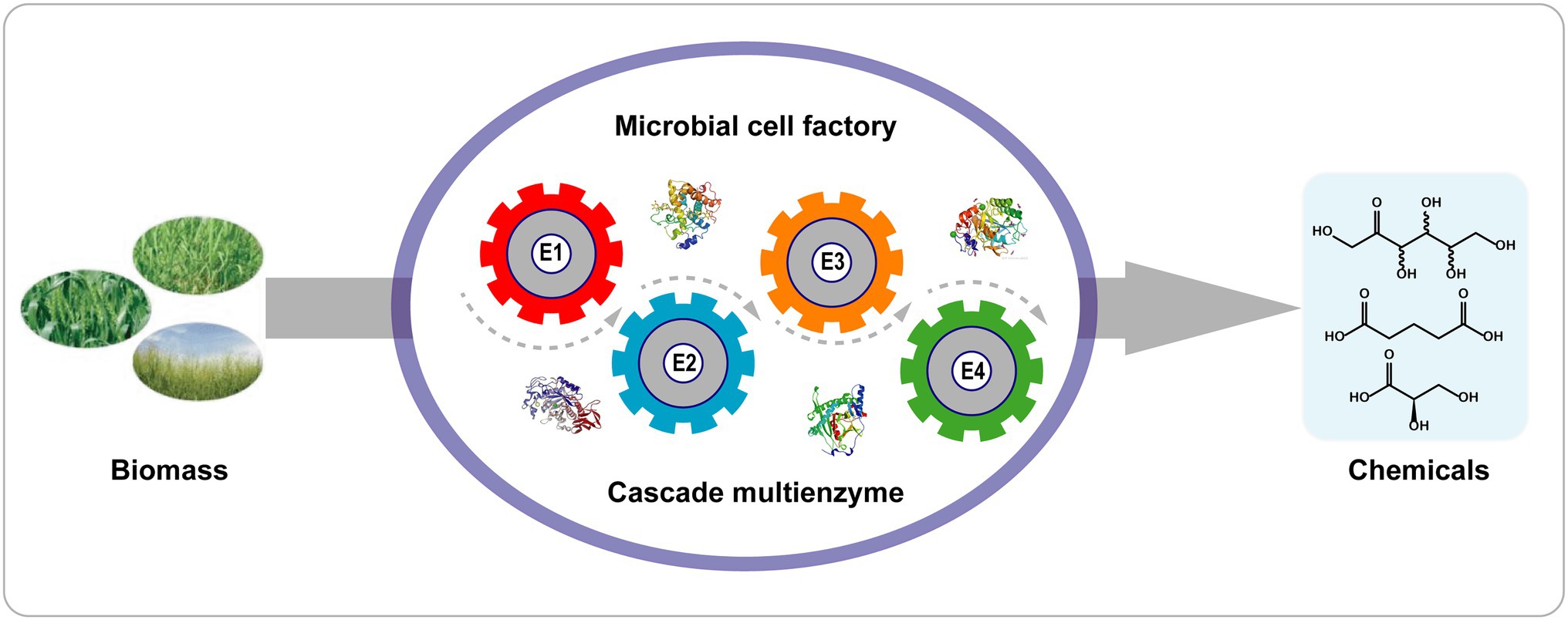
4. ábra . Kaszkád biotranszformáció egyetlen rekombináns sejttel. Az E1–E4 az 1–4. enzimeket jelöli.
A módosított Escherichia coli az egyik leggyakrabban használt organizmus a D-allulóz előállítására, tiszta háttere, gyors növekedési üteme, egyszerű tenyészete és stabil genetikája miatt.
Az Acidothermus cellulolyticus DAEáz-ját és D-glükóz izomerázát (GIáz) együtt expresszálták, hogy D-glükózból D-allulózt állítsanak elő ( Zhang et al., 2017 ).
Hasonlóképpen, a DAEáz-t és a xilóz izomerázt (XI) együtt expresszálták, hogy D-glükózt használva szubsztrátként D-allulózt állítsanak elő ( Chen et al., 2017 ).
A fenti két megközelítésben a D-glükózt először D-fruktózzá alakították a GIáz vagy a XI segítségével, majd a D-fruktózt azonnal izomerizálták D-allulózzá a DAEáz segítségével.
Bár az E. coli manipulálásával jó termelékenységet értek el , ami az endotoxinok és nem élelmiszeripari minőségű besorolásuk miatt az élelmiszeriparban nem volt alkalmazható, a DAEázt a mai napig sikeresen expresszálták számos élelmiszerbiztonságos törzsben, mint például a B. subtilis , a Saccharomyces cerevisiae és a Corynebacterium glutamicum ( Li et al., 2015b ; He et al., 2016 ). He és munkatársai (2016) a Clostridium scindens ATCC 35704 DTEázt jelenítették meg a B. subtilis spórák felületén, D-fruktózból D-allulóz előállításához. A DAEázt a CotZ C-terminálisához kapcsolták, és nagy hőstabilitást mutatott. Öt felhasználási ciklus után 60%-os aktivitás maradt fenn ( He et al., 2016 ).
Korábbi tanulmányunkban egy spórafelszíni megjelenítési technikát dolgoztunk ki D-allulóz előállítására D-glükózból. Ebben a megközelítésben a Thermus thermophilus XI. számú kulcsenzimjét és az A. tumefaciens DAEáz-ját biológiai, illetve kémiai módszerekkel immobilizálták az S. cerevisiae spórákon ( Li et al., 2015b ).
Ezenkívül több DAEázt és invertázt (INV) túlexpresszáltak a C. glutamicumban , és az algináttal immobilizált módosított sejteket kaszkádreakciónak vetették alá egy egylépéses, kétlépéses reakciórendszerben, hogy D-allulózt állítsanak elő nádmelaszból. 8 órás reakció után 61,2 g/L D-allulózt kaptak, ami a teljes monoszacharid 17,4%-át tette ki ( Yang et al., 2019 ).
Jelenleg az Izumoring stratégia a D-allulóz ipari előállításának fő módszere, amely SMB alkalmazásával érhető el. Ezenkívül az elválasztott D-fruktóz újra felhasználható D-allulóz előállítására. Röviden, az Izumoring stratégia a D-allulóz bioszintézisének legegyszerűbb és leggyakoribb módja. A termodinamikai egyensúly korlátai azonban szűk keresztmetszetet jelentenek, amely korlátozza a D-allulóz széles körű alkalmazását az élelmiszeriparban.
D-allulóz előállítása DHAP-függő aldolázok segítségével
Amint azt fentebb említettük, az Izumoring stratégián alapuló különféle megközelítéseket fejlesztettek ki a D-allulóz előállítására olcsó kiindulási anyagokból, például csicsókából, inulinból és mezőgazdasági maradványokból.
A DAEáz kulcsenzim által katalizált reakció azonban reverzibilis, és az átalakulási arány alacsony, ami a kapott D-allulóz magas árához vezet. Ezenkívül a D-allulóz szintézisében használt törzsek többsége nem GRAS mikroorganizmus.
Ezért rendkívül fontos egy költséghatékony és biztonságos platform létrehozása a D-allulóz tömegtermeléséhez.
Az aldolázok által katalizált sztereoszelektív aldoladdíciók alapvető eszközzé váltak a CC aszimmetrikus szintézisében ( Iturrate et al., 2010 ).
Az aldolázcsalád tagjai közül a DHAP-függő aldolázok a legelterjedtebbek számos olyan szénhidrátvegyület szintézisében, amelyeket nehéz szintetizálni hagyományos kémiai módszerekkel ( Bosshart et al., 2013 ).
Négyféle DHAP-függő aldoláz létezik: L-ramnulóz-1-foszfát aldoláz (RhaD), L-fukulóz-1-foszfát aldoláz (FucA), D-fruktóz-1,6-biszfoszfát aldoláz (FruA) és D-tagatóz-1,6-biszfoszfát aldoláz (TagA).
E négy aldoláz sztereoszelektív aldolreakciói kiegészítik egymást. Elméletileg négy ketózból álló készlet állítható elő, ha a DHAP-t donorként, és ugyanazt az aldehidet receptorként használjuk ( Gustavo, 2000 ).
Ezért a DHAP-függő aldolázok kiválóan alkalmasak különféle ritka cukrok (beleértve a D-allulózt, a D-szorbózt, a D-tagatózt és az L-fruktózt) szintézisére egyedi sztereoszelektivitásuk miatt ( 5. ábra ; Brovetto et al., 2011 ; Dai et al., 2021
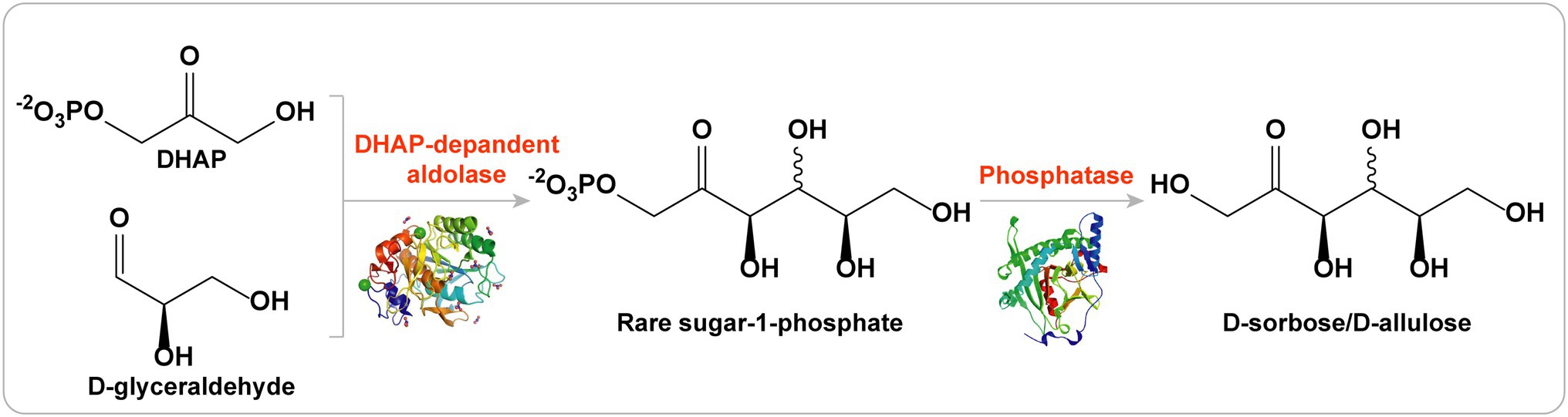
5. ábra . DHAP-függő aldolázok stratégiája D-allulóz előállítására.
D-allulóz bioszintézise RhaD aldolázzal, D-glicerinaldehid akceptorral
A DHAP-függő aldoláz stratégiák egyik fő hátránya, hogy a donor szubsztrát, a DHAP, nagyon drága és instabil a nagyméretű szintézishez ( Schümperli et al., 2007 ).
Azonban a DHAP szintézisének számos módja van enzimatikus stratégiákon keresztül . Például a DHAP előállítható dihidroxi-acetonon (DHA), glicerinen vagy glicerin-3-foszfáton keresztül , vagy metabolikus útvonalakon keresztül egy olcsó nyersanyagból, például glükózból vagy glicerinből ( 6. ábra ; Sánchez-Moreno et al., 2004 ; Li et al., 2012 , 2015a ; Wei et al. , 2015 ; Yang et al., 2015 , 2016 ).
A mesterségesen módosított E. coliban a RhaD által katalizált aldolreakció ritka cukrok (D-allulóz és D-szorbóz) szintézise céljából glükózt használtak szénforrásként, és folyamatosan adagoltak D-glicerinaldehidet a táptalajba. A fermentációs körülmények optimalizálását követően a D-allulóz és a D-szorbóz izolált hozama 0,21 mol/mol D-glicerinaldehid volt ( Wei et al., 2015 ). A D-allulózt és a D-szorbózt egy GRAS C. glutamicum törzsben állították elő glükóz és D-glicerinaldehid alapanyagokkal. A RhaD-t és fruktóz-1-foszfatázt (YqaB) hordozó rekombináns C. glutamicum törzsekről megállapították, hogy 19,5 g/l D-szorbózt és 13,4 g/l D-allulózt halmoznak fel fed szakaszos fermentációban ( Yang et al., 2015 ). Ezenkívül D-allulózt és D-szorbózt állítottak elő egy rekombináns E. coli törzsben, amely glicerinből fermentációval túltermeli az RhaD és YqaB aldolázt. 15 órás fermentáció után meghatároztuk a D-szorbóz (1,6 g/l) és a D-allulóz (1,23 g/l) koncentrációját a felülúszóban ( Li et al., 2015a ). Nemrégiben csoportunk egy hatékony rendszert konstruált a D-szorbóz és D-allulóz teljes sejtes kaszkádszintézisére glicerinből és D-glicerin-aldehidből, amely szakaszos biotranszformáció során 15,3 g/l D-szorbózt és 6,4 g/l D-allulózt eredményezett ( Chen et al., 2020a ).
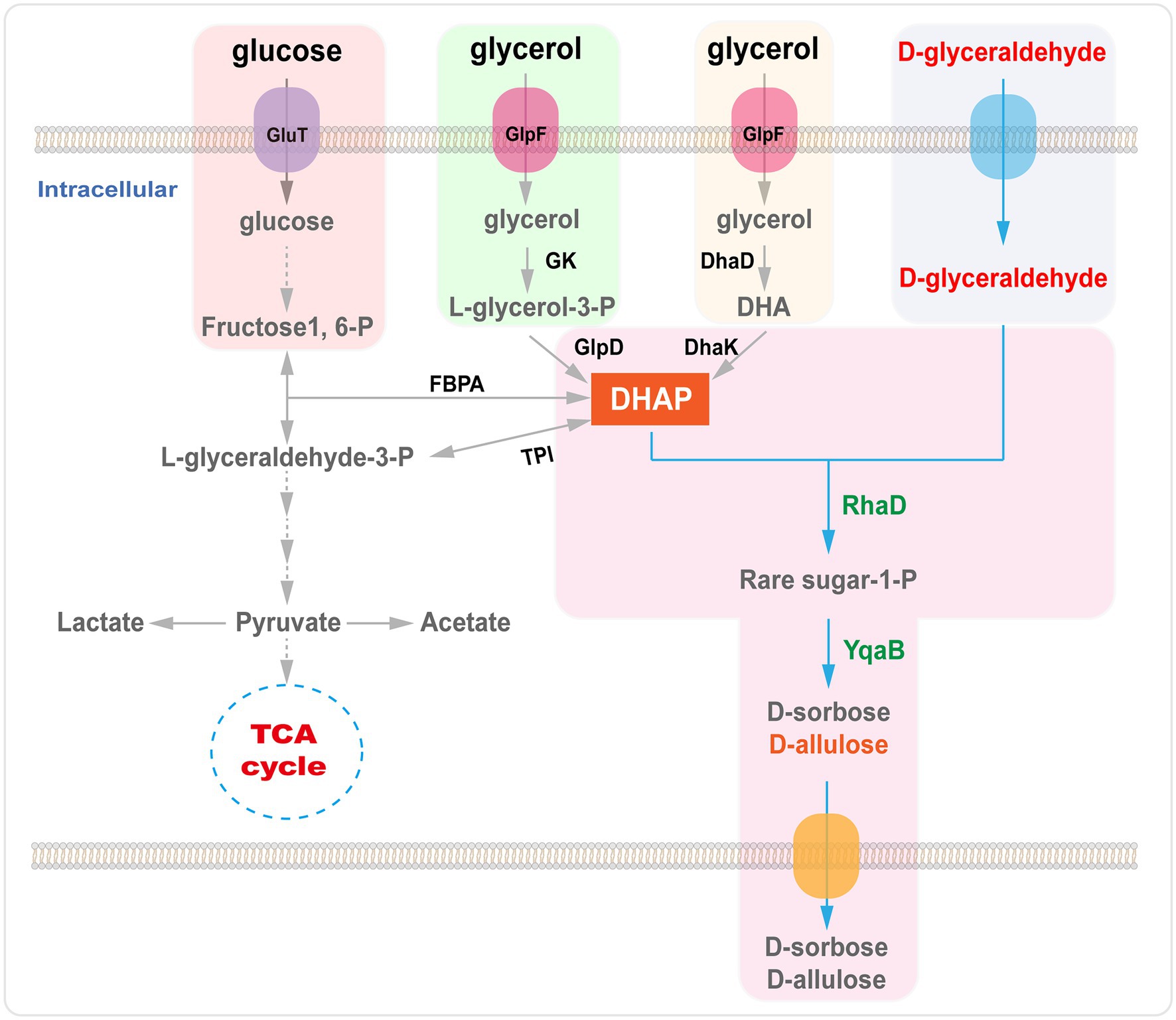
6. ábra . D-allulóz bioszintézise RhaD aldolázzal glicerinből vagy glükózból. GlpF, glicerin transzporter; GK, glicerin kináz; GlpD, glicerin-3-foszfát dehidrogenáz; DhaD, glicerin dehidrogenáz; DhaK, dihidroxi-aceton kinázok; FBPA, fruktóz-1,6-difoszfát aldoláz; TPI, trióz-foszfát izomeráz; RhaD, L-ramnulóz-1-foszfát aldoláz; YqaB, fruktóz-1-foszfatáz.
D-allulóz bioszintézise glicerinből, mint egyedüli szubsztrátból
Egy, az RhaD aldolázon alapuló mikroorganizmus-platform a D-allulóz szintéziséhez ígéretes lenne nagyszabású termelés szempontjából. A fő kihívást a donor szubsztrát DHAP és az akceptor molekula D-glicerin-aldehid magas költsége jelenti. Szerencsére megoldódott a DHAP felhalmozásának problémája az olcsó ipari melléktermék, a glicerin „zöld” szénforrásként való felhasználásával. Így a következő kérdés egy új enzim azonosítása, amely az alacsony értékű glicerint D-glicerin-aldehiddé alakítja. Szükség van egy olyan glicerin-dehidrogenázra vagy glicerin-oxidázra, amely hatékonyan képes katalizálni a glicerint D-glicerin-aldehiddé.
A probléma megoldása érdekében csoportunk egy platformot alkotott a D-szorbóz és a D-allulóz teljes sejtes kaszkádszintéziséhez, glicerint használva egyedüli szubsztrátként. Ebben a rendszerben ( 7. ábra ) a donor szubsztrát, a DHAP, a glicerin asszimilációs útvonalon keresztül keletkezik, az endogén DHAP pedig a glicerin-kináz (GK) és a glicerin-foszfát-oxidáz (GPO) túltermelésével termelődik . Az akceptor D-glicerin-aldehidet közvetlenül a glicerinből állítja elő a Streptomyces coelicolor A3 (AldO S.coe ) alditol-oxidáza. Ezután az RhaD katalizálja a DHAP és a D-glicerin-aldehid közötti aldolreakciót, így a megfelelő ketóz-1-foszfát keletkezik. Végül a D-szorbózt és a D-allulózt a foszfátcsoport YqaB foszfatázzal történő eltávolításával nyerik. Ezzel a rendszerrel a D-szorbóz és a D-allulóz termelési hozamai körülbelül 1,7-szeresére, illetve 1,2-szeresére nőttek a peroxidáz (Prx02 vagy KatE) túltermelése után, a H₂O₂ káros hatásainak kiküszöbölése érdekében . Összesen 7,9 g/l D-szorbózt és D-allulózt kaptunk glicerinből, 17,7%-os teljes konverziós arány mellett ( Chen et al., 2020b ).
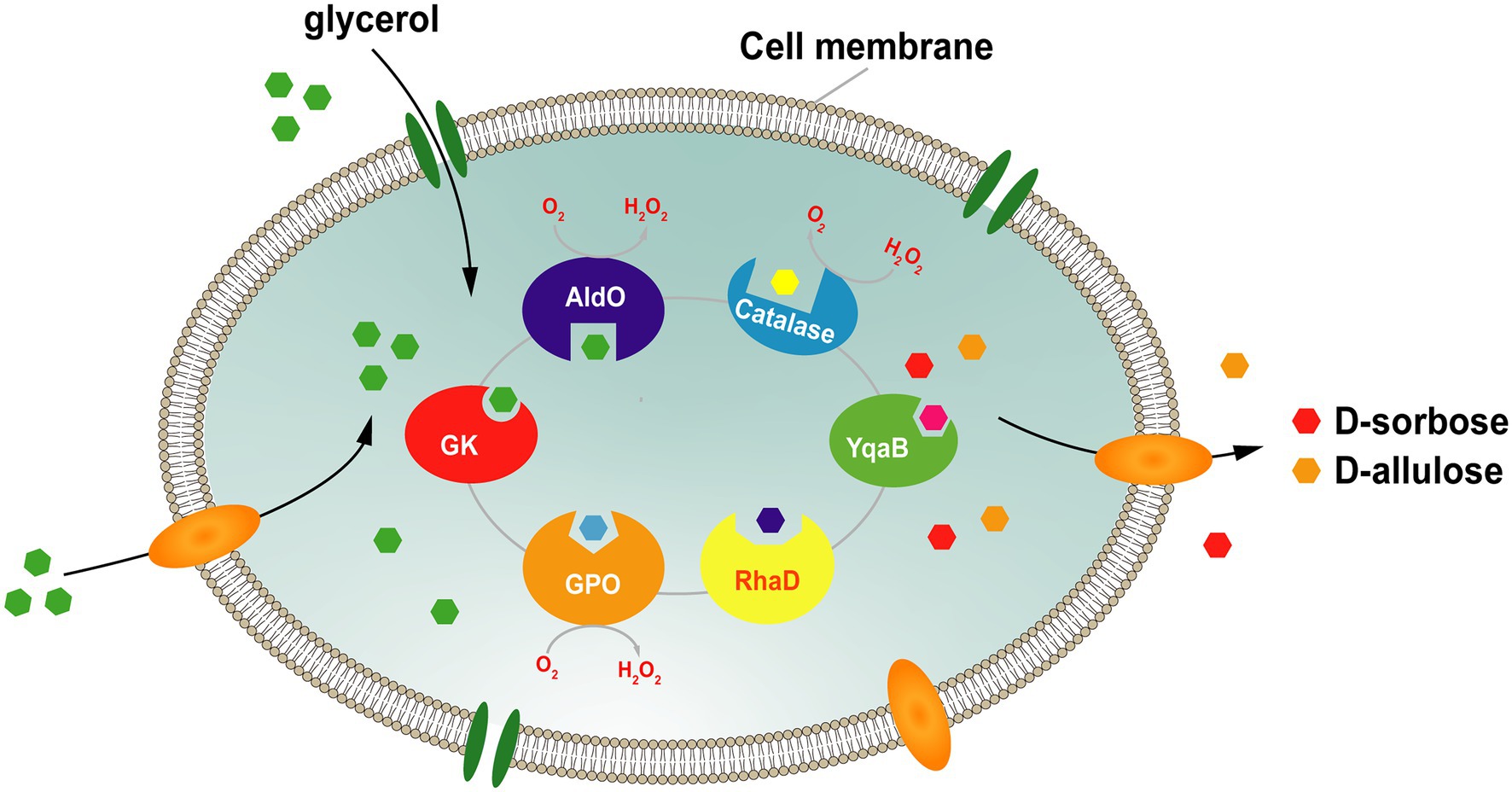
7. ábra . Stratégia a D-szorbóz és D-allulóz teljes sejtes szintézisére glicerinből. GK, glicerin-kináz; GPO, glicerin-oxidáz; AldO; alditol-oxidáz; RhaD, L-ramnulóz-1-foszfát-aldoláz; YqaB, fruktóz-1-foszfatáz.
Csoportunk egy egylépéses multienzimes rendszert is konstruált D-szorbóz és D-allulóz szintézisére glicerinből, mint egyetlen szénforrásból ( 8. ábra ). Itt a Shigella flexneriből származó PhoN savas foszfatázt (PhoN-Sf) vittünk a rendszerbe GK és YqaB helyett, amely katalizálta a glicerin foszforilációs reakcióját az első lépésben, és segített a ketóz-1-foszfát foszfátjának újrahasznosításában az utolsó lépésben, szabad ritkacukor molekulákat biztosítva. Az AldO S.coe-t vittük be a fenti multienzimes kaszkádba, hogy kizárólag a könnyen hozzáférhető glicerinből szintetizáljuk a D-szorbózt és a D-allulózt. Végül 14,8 g/L D-szorbózt és D-allulózt kaptunk glicerinből ( Li et al., 2020b ). A fenti stratégia egy nagyon hasznos és alacsony költségű megközelítést jelent különféle más ritka cukrok előállítására is. Egy szóval, a D-allulóz bioszintézise DHAP-függő aldolázokon alapuló fermentációval nagyon ígéretes. Jövedelmezőbb lenne egy DHAP-függő aldoláz stratégián alapuló sejtgyárat építeni a D-allulóz szintéziséhez.
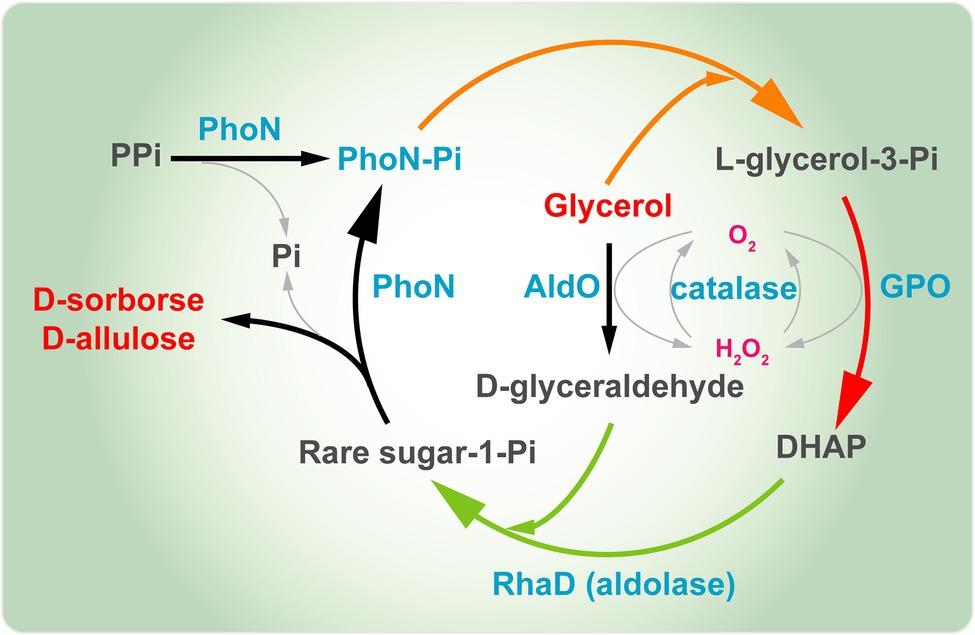
8. ábra . D-szorbóz és D-allulóz többenzimes szintézise glicerinből. PhoN, savas foszfatáz; GPO, glicerin-oxidáz; AldO; alditol-oxidáz; RhaD, L-ramnulóz-1-foszfát aldoláz; DHAP, dihidroxi-aceton-foszfát.
D-allulóz termodinamikavezérelt előállítása foszforiláció-defoszforiláció stratégián alapulva
Valójában a fenti stratégiák továbbra sem tudják kielégíteni az élelmiszeripar D-allulóz iránti igényét. Ezért érdekes egy új stratégia kidolgozása a D-allulóz előállítására, amely rendkívül hatékony és alacsony költségű zöld biogyártással jár. Nemrégiben You és munkatársai egy in vitro szintetikus enzimatikus bioszisztémát hoztak létre D-allulóz előállítására olcsó keményítőből, a „termodinamikailag vezérelt stratégia” alapján ( 9. ábra ). Ez az in vitro bioszisztéma öt fő enzimet foglalt magában, a reakciók a következőképpen mennek végbe: (1) a maltodextrint (a keményítő származékát) foszforilálták glükóz-1-foszfát (G1P) előállítására α-glükán-foszforiláz (αGP) segítségével, foszfát koszubsztráttal. (2) A G1P-t glükóz-6-foszfáttá (G6P) alakították, amit a foszfoglükomutáz (PGM) katalizált. (3) A G6P-t fruktóz-6-foszfáttá (F6P) alakították, amit a foszfoglükóz izomeráz (PGI) katalizált. (4) Az F6P epimerizációja D-allulóz-6-foszfát (A6P) előállítását eredményezte, amelyet a D-allulóz-6-foszfát 3-epimeráz (A6PE) katalizált. (5) Az A6P defoszforilációja D-allulóz és foszfát előállítását eredményezte, amelyet a D-allulóz-6-foszfát-foszfatáz (A6PP) katalizált. Ezenkívül a másik négy segédenzimet [izoamiláz (IA), 4-α-glükanotranszferáz (4GT), polifoszfát-glükokináz (PPGK) és glükóz-izomeráz (GI)] különböző időpontokban adtuk a reakciórendszerhez, hogy elérjük a maltodextrin teljes felhasználását a D-allulóz előállításához. Ebben a bioszisztémában az A6P defoszforilációjának Gibbs-energiája D-allulózzá -15,5 kJ/mol, ami irreverzibilis, ami azt jelzi, hogy a D-allulóz előállításához szükséges defoszforilációs lépés termodinamikailag kedvező és egyirányú, hogy a teljes reakciót a teljesség felé terelje. A reakciókörülmények optimalizálása után a D-allulóz termelési hozama 10 és 50 g/l keményítőből elérték a 88,2%-ot, illetve a 79,2%-ot ( Li et al., 2021 ). Összességében ez a termodinamika-vezérelt stratégia ígéretes alternatívát kínál a D-allulóz költséghatékony előállítására. Mivel a rendszer több enzimet tartalmaz, és az enzimtisztítás lépései nehézkesek, a fenti stratégia továbbra sem tudja kielégíteni az iparosítás igényeit.
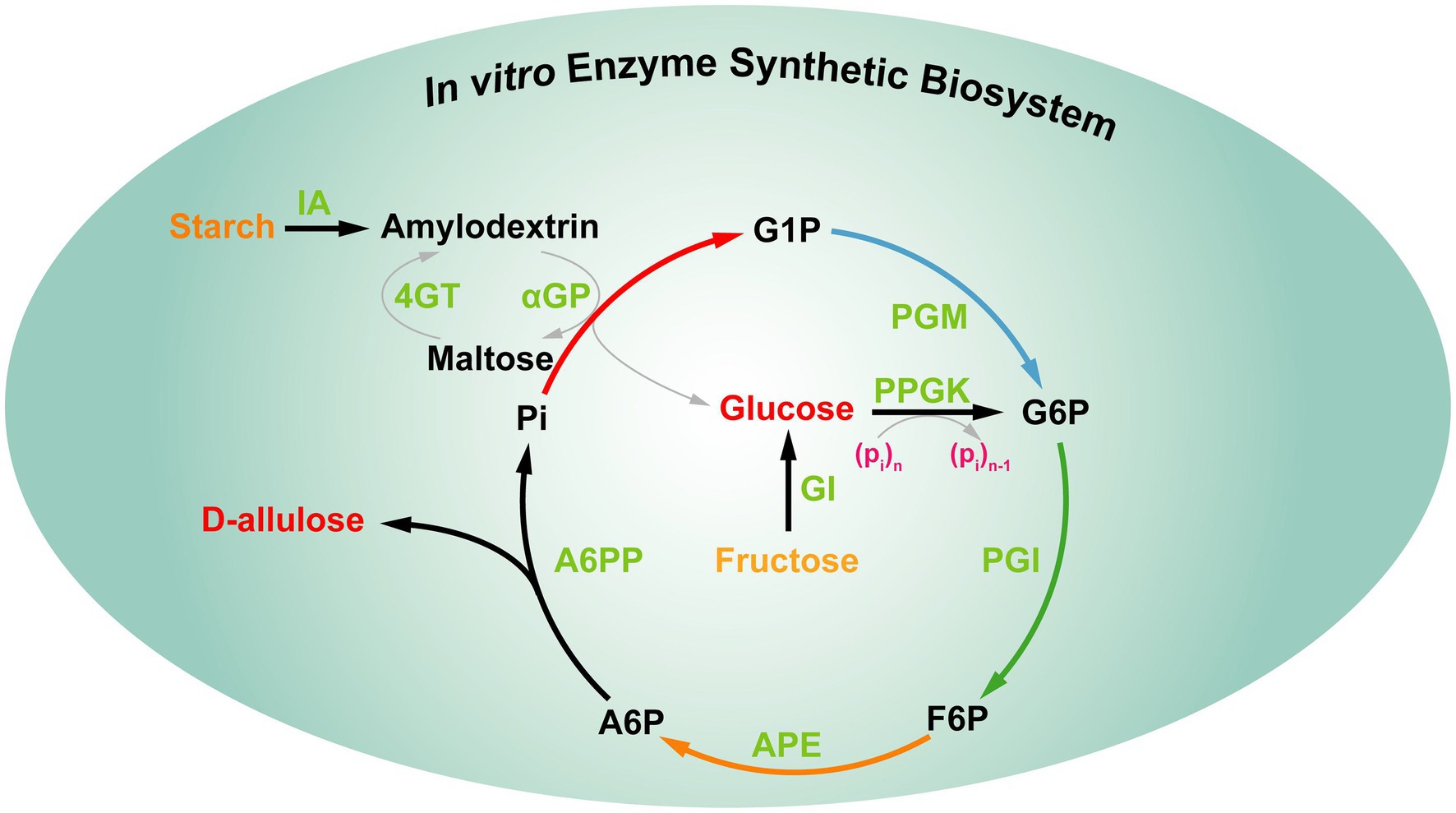
9. ábra . D-allulóz in vitro többenzimes szintézise keményítőből. IA, izoamiláz; 4GT, 4-α-glükanotranszferáz; αGP, α-glükán-foszforiláz; PGM, foszfoglükomutáz; PPGK, polifoszfát-glükokináz; PGI, foszfoglükóz-izomeráz; GI, glükóz-izomeráz; APE, D-allulóz-6-foszfát-3-epimeráz; A6PP, D-allulóz-6-foszfát-foszfatáz; G1P, glükóz-1-foszfát; G6P, glükóz-6-foszfát; F6P, fruktóz-6-foszfát; A6P, D-allulóz-6-foszfát.
Összefoglalás és jövőbeli kilátások
A D-allulóz előállításának leggyakoribb módszere az Izumoring stratégián alapul, amelyet a termodinamikai egyensúly korlátoz, ami alacsony konverziós arányt és viszonylag magas szintetikus költségeket eredményez.
Ezért új és robusztus DTEáz családba tartozó enzimeket kell felfedezni. Továbbá az irány-evolúció stratégiája. (Zhang et al., 2016a; Zhu et al., 2019a) vagy enzim immobilizálás (Ran et al., 2019; Wong et al., 2020)
A DTEáz katalitikus teljesítményének javítása nagyon hasznos lesz. A DHAP-függő aldoláz stratégiához a D-allulóz és a D-szorbóz egyidejűleg keletkezik RhaD-val, D-glicerol-aldehid akceptorral. Az aldolázok sztereoszelektivitásának javítása érdekében számos fejlett technikát és módszert alkalmaznak, beleértve az irányított evolúciót is. (d’Oelsnitz and Ellington, 2018; Shepelin et al., 2018; Currin et al., 2021), nagy áteresztőképességű szűrőtechnikák (Ung et al., 2018; Rienzo et al., 2021), és racionális mérnöki (Damborsky and Brezovsky, 2014; Windle et al., 2014; Kim et al., 2020), alkalmazható.
Nyilvánvaló, hogy a termodinamika-vezérelt stratégia ígéretes módszer a D-allulóz előállítására. A jövő egyik iránya egy ilyen megközelítést alkalmazó mikrobiális sejtgyár felépítése lenne. Számos anyagcsere-eszköz, például a CRISPR/Cas9, hozzájárulhat a D-allulóz ipari előállításához. (Wu et al., 2019; Nishida and Kondo, 2021), önösszeszerelés (Liu et al., 2019; Lange and Polizzi, 2021), és dinamikus szabályozás (Hartline et al., 2021; Zhu et al., 2021).
Remélhetőleg a D-allulóz a közeljövőben megfizethetővé válik a hétköznapi fogyasztók számára is.
Szerzők hozzájárulásai
ZC végezte az adatok feldolgozását és írását – az eredeti vázlat elkészítését. X-DG részt vett a vizualizációban, a vizsgálatban és a felügyeletben. ZL végezte az írást – a lektorálást és a szerkesztést. Minden szerző hozzájárult a cikkhez, és jóváhagyta a benyújtott verziót.
Finanszírozás
Ezt a munkát a Kínai Nemzeti Természettudományi Alapítvány (számok: 32171475, 31971216), a Kínai Posztdoktori Tudományos Alapítvány (szám: 2021M691285), a Shandong Tartományi Nagy Tudományos és Technológiai Innovációs Projekt (szám: 2019JZZY011006), a Jiangsu Tartományi Természettudományi Alapítvány (szám: BK20210465), valamint a Szakterületi Tehetségek Egyetemi Bevezetésének Programja (szám: 111-2-06) támogatta.
Érdekellentét
A szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.
Kiadói megjegyzés
A cikkben kifejtett összes állítás kizárólag a szerzők állítása, és nem feltétlenül tükrözi kapcsolt szervezeteik, a kiadó, a szerkesztők és a lektorok állításait. A cikkben értékelt termékekre, illetve a gyártó által tett állításokra a kiadó nem vállal garanciát és nem is támogatja azokat.
Köszönetnyilvánítás
Köszönjük Zhou Xiaomannak (Jiangnan Egyetem) a rajzot.
Referenciák
Basu, A., Basu, R., Shah, P., Vella, A., Johnson, C. M., Jensen, M., et al. (2001). A 2-es típusú cukorbetegség károsítja a glükóz splanchnikus felvételét, de nem változtatja meg a bélből származó glükóz felszívódását az enterális glükóztáplálás során: további bizonyíték a máj glükokináz aktivitásának hibájára. Diabetes 50, 1351–1362. doi: 10.2337/diabetes.50.6.1351
Bilal, M., Iqbal, H. M. N., Hu, H., Wang, W., and Zhang, X. (2018). Metabolic engineering pathways for rare sugars biosynthesis, physiological functionalities, and applications-a review. Crit. Rev. Food Sci. Nutr. 58, 2768–2778. doi: 10.1080/10408398.2017.1341385
Bosshart, A., Panke, S., and Bechtold, M. (2013). Systematic optimization of interface interactions increases the thermostability of a multimeric enzyme. Angew. Chem. Int. Ed. Engl. 52, 9673–9676. doi: 10.1002/anie.201304141
Brovetto, M., Gamenara, D., Méndez, P. S., and Seoane, G. A. (2011). C-C bond-forming lyases in organic synthesis. Chem. Rev. 111, 4346–4403. doi: 10.1021/cr100299p
Castro-Muñoz, R., Correa-Delgado, M., Córdova-Almeida, R., Lara-Nava, D., Chávez-Muñoz, M., Velásquez-Chávez, V. F., et al. (2022). Natural sweeteners: sources, extraction and current uses in foods and food industries. Food Chem. 370:130991. doi: 10.1016/j.foodchem.2021.130991
Chen, J., Huang, W., Zhang, T., Lu, M., and Jiang, B. (2019). Anti-obesity potential of rare sugar D-psicose by regulating lipid metabolism in rats. Food Funct. 10, 2417–2425. doi: 10.1039/c8fo01089g
Chen, Z., Li, Z., Li, F., Wang, N., and Gao, X.-D. (2020b). Characterization of alditol oxidase from Streptomyces coelicolor and its application in the production of rare sugars. Bioorg. Med. Chem. 28:115464. doi: 10.1016/j.bmc.2020.115464
Chen, Z., Li, Z., Li, F., Wang, M., Wang, N., and Gao, X.-D. (2020a). Cascade synthesis of rare ketoses by whole cells based on L-rhamnulose-1-phosphate aldolase. Enzym. Microb. Technol. 133:109456. doi: 10.1016/j.enzmictec.2019.109456
Chen, X., Wang, W., Xu, J., Yuan, Z., Yuan, T., Zhang, Y., et al. (2017). Production of D-psicose from D-glucose by co-expression of D-psicose 3-epimerase and xylose isomerase. Enzym. Microb. Technol. 105, 18–23. doi: 10.1016/j.enzmictec.2017.06.003
Chen, J., Zhu, Y., Fu, G., Song, Y., Jin, Z., Sun, Y., et al. (2016). High-level intra- and extra-cellular production of D-psicose 3-epimerase via a modified xylose-inducible expression system in Bacillus subtilis. J. Ind. Microbiol. Biotechnol. 43, 1577–1591. doi: 10.1007/s10295-016-1819-6
Chung, Y. M., Hyun Lee, J., Youl Kim, D., Hwang, S. H., Hong, Y. H., Kim, S. B., et al. (2012). Dietary D-psicose reduced visceral fat mass in high-fat diet-induced obese rats. J. Food Sci. 77, H53–H58. doi: 10.1111/j.1750-3841.2011.02571.x
Currin, A., Parker, S., Robinson, C. J., Takano, E., Scrutton, N. S., and Breitling, R. (2021). The evolving art of creating genetic diversity: From directed evolution to synthetic biology. Biotechnol. Adv. 50:107762. doi: 10.1016/j.biotechadv.2021.107762
Dai, Y., Zhang, J., Jiang, B., Zhang, T., and Chen, J. (2021). New strategy for rare sugars biosynthesis: Aldol reactions using dihydroxyacetone phosphate (DHAP)-dependent aldolases. Food Biosci. 44:101377. doi: 10.1016/j.fbio.2021.101377
Damborsky, J., and Brezovsky, J. (2014). Computational tools for designing and engineering enzymes. Curr. Opin. Chem. Biol. 19, 8–16. doi: 10.1016/j.cbpa.2013.12.003
d’Oelsnitz, S., and Ellington, A. (2018). Continuous directed evolution for strain and protein engineering. Curr. Opin. Biotechnol. 53, 158–163. doi: 10.1016/j.copbio.2017.12.020
Doner, L. W. (1979). Isomerization of D-fructose by base: liquid-chromatographic evaluation and the isolation of D-psicose. Carbohydr. Res. 70, 209–216. doi: 10.1016/S0008-6215(00)87101-3
Granström, T. B., Takata, G., Tokuda, M., and Izumori, K. (2004). Izumoring: A novel and complete strategy for bioproduction of rare sugars. J. Biosci. Bioeng. 97, 89–94. doi: 10.1016/S1389-1723(04)70173-5
Grassi, B., Onetto, M. T., Zapata, Y., Jofré, P., and Echeverría, G. (2021). Lower versus standard sucrose dose for treating hypoglycemia in patients with type 1 diabetes mellitus in therapy with predictive low glucose suspend (PLGS) augmented insulin pumps: a randomized crossover trial in Santiago, Chile. Diabetes Metab. Syndr. Clin. Res. Rev. 15, 695–701. doi: 10.1016/j.dsx.2021.03.017
Guerrero-Wyss, M., Durán Agüero, S., and Angarita Dávila, L. (2018). D-Tagatose is a promising sweetener to control glycaemia: A new functional food. Biomed. Res. Int. 2018:8718053. doi: 10.1155/2018/8718053
Gustavo, S. (2000). Enzymatic C-C bond-forming reactions in organic synthesis. Curr. Org. Chem. 4, 283–304. doi: 10.2174/1385272003376283
Han, Y., Han, H. J., Kim, A. H., Choi, J. Y., Cho, S. J., Park, Y. B., et al. (2016). D-Allulose supplementation normalized the body weight and fat-pad mass in diet-induced obese mice via the regulation of lipid metabolism under isocaloric fed condition. Mol. Nutr. Food Res. 60, 1695–1706. doi: 10.1002/mnfr.201500771
Hartline, C. J., Schmitz, A. C., Han, Y., and Zhang, F. (2021). Dynamic control in metabolic engineering: theories, tools, and applications. Metab. Eng. 63, 126–140. doi: 10.1016/j.ymben.2020.08.015
Hayakawa, M., Hira, T., Nakamura, M., Iida, T., Kishimoto, Y., and Hara, H. (2018). Secretion of GLP-1 but not GIP is potently stimulated by luminal D-Allulose (D-Psicose) in rats. Biochem. Biophys. Res. Commun. 496, 898–903. doi: 10.1016/j.bbrc.2018.01.128
Hayashi, N., Iida, T., Yamada, T., Okuma, K., Takehara, I., Yamamoto, T., et al. (2010). Study on the postprandial blood glucose suppression effect of D-psicose in borderline diabetes and the safety of long-term ingestion by normal human subjects. Biosci. Biotechnol. Biochem. 74, 510–519. doi: 10.1271/bbb.90707
He, W., Jiang, B., Mu, W., and Zhang, T. (2016). Production of D-Allulose with D-psicose 3-epimerase expressed and displayed on the surface of Bacillus subtilis spores. J. Agric. Food Chem. 64, 7201–7207. doi: 10.1021/acs.jafc.6b03347
Herling, A. W., Burger, H.-J., Schwab, D., Hemmerle, H., Below, P., and Schubert, G. (1998). Pharmacodynamic profile of a novel inhibitor of the hepatic glucose-6-phosphatase system. Am. J. Phys. 274, G1087–G1093. doi: 10.2337/diabetes.47.5.85
Hoshikawa, H., Kamitori, K., Indo, K., Mori, T., Kamata, M., Takahashi, T., et al. (2018). Combined treatment with D-allose, docetaxel and radiation inhibits the tumor growth in an in vivo model of head and neck cancer. Oncol. Lett. 15, 3422–3428. doi: 10.3892/ol.2018.7787
Hossain, A., Yamaguchi, F., Matsuo, T., Tsukamoto, I., Toyoda, Y., Ogawa, M., et al. (2015). Rare sugar D-allulose: potential role and therapeutic monitoring in maintaining obesity and type 2 diabetes mellitus. Pharmacol. Therapeut. 155, 49–59. doi: 10.1016/j.pharmthera.2015.08.004
Iida, T., Kishimoto, Y., Yoshikawa, Y., Hayashi, N., Okuma, K., Tohi, M., et al. (2008). Acute D-psicose administration decreases the glycemic responses to an oral maltodextrin tolerance test in normal adults. J. Nutr. Sci. Vitaminol. 54, 511–514. doi: 10.3177/jnsv.54.511
Itoh, H., Sato, T., and Izumori, K. (1995). Preparation of D-psicose from D-fructose by immobilized D-tagatose 3-epimerase. J. Ferment. Bioeng. 79, 184–185. doi: 10.1016/0922-338X(95)94091-5
Iturrate, L., Sánchez-Moreno, I., Oroz-Guinea, I., Pérez-Gil, J., and García-Junceda, E. (2010). Preparation and characterization of a bifunctional aldolase/kinase enzyme: a more efficient biocatalyst for C-C bond formation. Chemistry 16, 4018–4030. doi: 10.1002/chem.200903096
Iwasaki, Y., Sendo, M., Dezaki, K., Hira, T., Sato, T., Nakata, M., et al. (2018). GLP-1 release and vagal afferent activation mediate the beneficial metabolic and chronotherapeutic effects of D-allulose. Nat. Commun. 9:113. doi: 10.1038/s41467-017-02488-y
Izumori, K. (2006). Izumoring: a strategy for bioproduction of all hexoses. J. Biotechnol. 124, 717–722. doi: 10.1016/j.jbiotec.2006.04.016
Jia, M., Mu, W., Chu, F., Zhang, X., Jiang, B., Zhou, L. L., et al. (2014). A D-psicose 3-epimerase with neutral pH optimum from Clostridium bolteae for D-psicose production: cloning, expression, purification, and characterization. Appl. Microbiol. Biotechnol. 98, 717–725. doi: 10.1007/s00253-013-4924-8
Kanasaki, A., Jiang, Z., Mizokami, T., Shirouchi, B., Iida, T., Nagata, Y., et al. (2019). D-allulose alters serum cholesterol levels partly via reducing serum pcsk9 levels in hamsters. Atherosclerosis 287:e201. doi: 10.1016/j.atherosclerosis.2019.06.611
Khan, S. A., Verma, P., Ur Rahman, L., and Parasharami, V. A. (2021). “Chapter 13 – Exploration of biotechnological studies in low-calorie sweetener Stevia rebaudiana: present and future prospects,” in Medicinal and Aromatic Plants: Expanding Their Horizons Through Omics. eds. T. Aftab and K. R. Hakeem (Academic Press), 289–324.
Kim, H.-J., Hyun, E.-K., Kim, Y.-S., Lee, Y.-J., and Oh, D.-K. (2006). Characterization of an agrobacterium tumefaciens D-psicose 3-epimerase that converts D-fructose to D-psicose. Appl. Environ. Microbiol. 72, 981–985. doi: 10.1128/AEM.72.2.981-985.2006
Kim, S.-E., Kim, S. J., Kim, H.-J., and Sung, M.-K. (2017). D-Psicose, a sugar substitute, suppresses body fat deposition by altering networks of inflammatory response and lipid metabolism in C57BL/6J-ob/ob mice. J. Funct. Foods 28, 265–274. doi: 10.1016/j.jff.2016.11.029
Kim, T., Stogios, P. J., Khusnutdinova, A. N., Nemr, K., Skarina, T., Flick, R., et al. (2020). Rational engineering of 2-deoxyribose-5-phosphate aldolases for the biosynthesis of (R)-1,3-butanediol. J. Biol. Chem. 295, 597–609. doi: 10.1074/jbc.RA119.011363
Kimura, T., Kanasaki, A., Hayashi, N., Yamada, T., Iida, T., Nagata, Y., et al. (2017). D-Allulose enhances postprandial fat oxidation in healthy humans. Nutrition 43–44, 16–20. doi: 10.1016/j.nut.2017.06.007
Lai, W. T., Khong, N. M. H., Lim, S. S., Hee, Y. Y., Sim, B. I., Lau, K. Y., et al. (2017). A review: modified agricultural by-products for the development and fortification of food products and nutraceuticals. Trends Food Sci. Technol. 59, 148–160. doi: 10.1016/j.tifs.2016.11.014
Lange, O. J., and Polizzi, K. M. (2021). Click it or stick it: covalent and non-covalent methods for protein-self assembly. Curr. Opin. Syst. Biol. 28:100374. doi: 10.1016/j.coisb.2021.100374
Lee, D., Han, Y., Kwon, E. Y., and Choi, M. S. (2020). D-allulose ameliorates metabolic dysfunction in C57BL/KsJ-db/db mice. Molecules 25:3656. doi: 10.3390/molecules25163656
Li, A., Cai, L., Chen, Z., Wang, M., Wang, N., Nakanishi, H., et al. (2017). Recent advances in the synthesis of rare sugars using DHAP-dependent aldolases. Carbohydr. Res. 452, 108–115. doi: 10.1016/j.carres.2017.10.009
Li, Z., Cai, L., Wei, M., and Wang, P. G. (2012). One-pot four-enzyme synthesis of ketoses with fructose 1,6-bisphosphate aldolases from Staphylococcus carnosus and rabbit muscle. Carbohydr. Res. 357, 143–146. doi: 10.1016/j.carres.2012.05.007
Li, Z., He, B., Gao, Y., and Cai, L. (2015a). Synthesis of D-sorbose and D-psicose by recombinant Escherichia coli. J. Carbohydr. Chem. 34, 349–357. doi: 10.1080/07328303.2015.1068794
Li, Z., Li, F., Cai, L., Chen, Z., Qin, L., and Gao, X.-D. (2020b). One-pot multienzyme synthesis of rare ketoses from glycerol. J. Agric. Food Chem. 68, 1347–1353. doi: 10.1021/acs.jafc.9b06748
Li, Z., Li, Y., Duan, S., Liu, J., Yuan, P., Nakanishi, H., et al. (2015b). Bioconversion of D-glucose to D-psicose with immobilized D-xylose isomerase and D-psicose 3-epimerase on Saccharomyces cerevisiae spores. J. Ind. Microbiol. 42, 1117–1128. doi: 10.1007/s10295-015-1631-8
Li, C., Lin, J., Guo, Q., Zhang, C., Du, K., Lin, H., et al. (2018). D-Psicose 3-epimerase secretory overexpression, immobilization, and D-psicose biotransformation, separation and crystallization. J. Chem. Technol. Biotechnol. 93, 350–357. doi: 10.1002/jctb.5360
Li, Y., Shi, T., Han, P., and You, C. (2021). Thermodynamics-driven production of value-added D-allulose from inexpensive starch by an in vitro enzymatic synthetic biosystem. ACS Catal. 11, 5088–5099. doi: 10.1021/acscatal.0c05718
Li, W., Zhu, Y., Jiang, X., Zhang, W., Guang, C., and Mu, W. (2020a). One-pot production of D-allulose from inulin by a novel identified thermostable exoinulinase from Aspergillus piperis and Dorea sp. D-allulose 3-epimerase. Process Biochem. 99, 87–95. doi: 10.1016/j.procbio.2020.08.021
Lim, B. C., Kim, H. J., and Oh, D. K. (2009). A stable immobilized D-psicose 3-epimerase for the production of D-psicose in the presence of borate. Process Biochem. 44, 822–828. doi: 10.1016/j.procbio.2009.03.017
Liu, S., Ammirati, M. J., Song, X., Knafels, J. D., Zhang, J., Greasley, S. E., et al. (2012). Insights into mechanism of glucokinase activation: observation of multiple distinct protein conformations. J. Biol. Chem. 287, 13598–13610. doi: 10.1074/jbc.M111.274126
Liu, Z., Cao, S., Liu, M., Kang, W., and Xia, J. (2019). Self-assembled multienzyme nanostructures on synthetic protein scaffolds. ACS Nano 13, 11343–11352. doi: 10.1021/acsnano.9b04554
Lloyd, D. J., St Jean, D. J., Kurzeja, R. J., Wahl, R. C., Michelsen, K., Cupples, R., et al. (2013). Antidiabetic effects of glucokinase regulatory protein small-molecule disruptors. Nature 504, 437–440. doi: 10.1038/nature12724
Matsuo, T., and Izumori, K. (2009). D-Psicose inhibits intestinal alpha-glucosidase and suppresses the glycemic response after ingestion of carbohydrates in rats. J. Clin. Biochem. Nutr. 45, 202–206. doi: 10.3164/jcbn.09-36
Matsuo, T., Suzuki, H., Hashiguchi, M., and Izumori, K. (2002). D-psicose is a rare sugar that provides no energy to growing rats. J. Nutr. Sci. Vitaminol. 48, 77–80. doi: 10.3177/jnsv.48.77
McDonald, E. J. (1967). A new synthesis of D-psicose (D-ribo-hexulose). Carbohydr. Res. 5, 106–108. doi: 10.1016/0008-6215(67)85014-6
Moller, D., and Berger, J. (2003). Role of PPARs in the regulation of obesity-related insulin sensitivity and inflammation. Int. J. Obes. 27, S17–S21. doi: 10.1038/sj.ijo.0802494
Nagata, Y., Kanasaki, A., Tamaru, S., and Tanaka, K. (2015). D-psicose, an epimer of D-fructose, favorably alters lipid metabolism in Sprague–Dawley rats. J. Agric. Food Chem. 63, 3168–3176. doi: 10.1021/jf502535p
Narayan Patel, S., Singh, V., Sharma, M., Sangwan, R. S., Singhal, N. K., and Singh, S. P. (2018). Development of a thermo-stable and recyclable magnetic nanobiocatalyst for bioprocessing of fruit processing residues and D-allulose synthesis. Bioresour. Technol. 247, 633–639. doi: 10.1016/j.biortech.2017.09.112
Natsume, Y., Yamada, T., Iida, T., Ozaki, N., Gou, Y., Oshida, Y., et al. (2021). Investigation of D-allulose effects on high-sucrose diet-induced insulin resistance via hyperinsulinemic-euglycemic clamps in rats. Heliyon 7:e08013. doi: 10.1016/j.heliyon.2021.e08013
Nishida, K., and Kondo, A. (2021). CRISPR-derived genome editing technologies for metabolic engineering. Metab. Eng. 63, 141–147. doi: 10.1016/j.ymben.2020.12.002
Ochiai, M., Onishi, K., Yamada, T., Iida, T., and Matsuo, T. (2014). D-psicose increases energy expenditure and decreases body fat accumulation in rats fed a high-sucrose diet. Int. J. Food Sci. Nutr. 65, 245–250. doi: 10.3109/09637486.2013.845653
Park, S. Y., and Yoon, K. Y. (2015). Enzymatic production of soluble dietary fiber from the cellulose fraction of Chinese cabbage waste and potential use as a functional food source. Food Sci. Biotechnol. 24, 529–535. doi: 10.1007/s10068-015-0069-0
Pfefferkorn, J. A. (2013). Strategies for the design of hepatoselective glucokinase activators to treat type 2 diabetes. Expert. Opin. Drug. Discov. 8, 319–330. doi: 10.1517/17460441.2013.748744
Qi, Z., Zhu, Z., Wang, J.-W., Li, S., Guo, Q., Xu, P., et al. (2017). Biochemical analysis and the preliminary crystallographic characterization of D-tagatose 3-epimerase from Rhodobacter sphaeroides. Microb. Cell Factories 16:193. doi: 10.1186/s12934-017-0808-4
Ran, G., Tan, D., Zhao, J., Fan, F., Zhang, Q., Wu, X., et al. (2019). Functionalized polyhydroxyalkanoate nano-beads as a stable biocatalyst for cost-effective production of the rare sugar D-allulose. Bioresour. Technol. 289:121673. doi: 10.1016/j.biortech.2019.121673
Rienzo, M., Jackson, S. J., Chao, L. K., Leaf, T., Schmidt, T. J., Navidi, A. H., et al. (2021). High-throughput screening for high-efficiency small-molecule biosynthesis. Metab. Eng. 63, 102–125. doi: 10.1016/j.ymben.2020.09.004
Sánchez-Moreno, I., Francisco García-García, J., Bastida, A., and García-Junceda, E. (2004). Multienzyme system for dihydroxyacetone phosphate-dependent aldolase catalyzed C–C bond formation from dihydroxyacetone. Chem. Commun. 35, 1634–1635. doi: 10.1002/chin.200447149
Schümperli, M., Pellaux, R., and Panke, S. (2007). Chemical and enzymatic routes to dihydroxyacetone phosphate. Appl. Microbiol. Biotechnol. 75, 33–45. doi: 10.1007/s00253-007-0882-3
Shepelin, D., Hansen, A. S. L., Lennen, R., Luo, H., and Herrgård, M. J. (2018). Selecting the best: evolutionary engineering of chemical production in microbes. Genes 9:249. doi: 10.3390/genes9050249
Shintani, T., Sakoguchi, H., Yoshihara, A., Izumori, K., and Sato, M. (2017a). D-Allulose, a stereoisomer of D-fructose, extends Caenorhabditis elegans lifespan through a dietary restriction mechanism: a new candidate dietary restriction mimetic. Biochem. Biophys. Res. Commun. 493, 1528–1533. doi: 10.1016/j.bbrc.2017.09.147
Shintani, T., Yamada, T., Hayashi, N., Iida, T., Nagata, Y., Ozaki, N., et al. (2017b). Rare sugar syrup containing D-allulose but not high-fructose corn syrup maintains glucose tolerance and insulin sensitivity partly via hepatic glucokinase translocation in Wistar rats. J. Agric. Food Chem. 65, 2888–2894. doi: 10.1021/acs.jafc.6b05627
Song, Y., Nguyen, Q. A., Wi, S. G., Yang, J., and Bae, H. J. (2016). Strategy for dual production of bioethanol and D-psicose as value-added products from cruciferous vegetable residue. Bioresour. Technol. 223, 34–39. doi: 10.1016/j.biortech.2016.10.021
Song, Y., Oh, C., and Bae, H.-J. (2017). Simultaneous production of bioethanol and value-added D-psicose from Jerusalem artichoke (Helianthus tuberosus L.) tubers. Bioresour. Technol. 244, 1068–1072. doi: 10.1016/j.biortech.2017.08.079
Su, L., Sun, F., Liu, Z., Zhang, K., and Wu, J. (2018). Highly efficient production of clostridium cellulolyticum H10 D-psicose 3-epimerase in Bacillus subtilis and use of these cells to produce D-psicose. Microb. Cell Factories 17:188. doi: 10.1186/s12934-018-1037-1
Suna, S., Yamaguchi, F., Kimura, S., Tokuda, M., and Jitsunari, F. (2007). Preventive effect of D-psicose, one of rare ketohexoses, on di-(2-ethylhexyl) phthalate (DEHP)-induced testicular injury in rat. Toxicol. Lett. 173, 107–117. doi: 10.1016/j.toxlet.2007.06.015
Takata, M. K., Yamaguchi, F., Nakanose, K., Watanabe, Y., Hatano, N., Tsukamoto, I., et al. (2005). Neuroprotective effect of D-psicose on 6-hydroxydopamine-induced apoptosis in rat pheochromocytoma (PC12) cells. J. Biosci. Bioeng. 100, 511–516. doi: 10.1263/jbb.100.511
Toyoda, Y., Mori, S., Umemura, N., Futamura, N., Inoue, H., Hata, T., et al. (2010). Suppression of blood glucose levels by D-psicose in glucose tolerance test in diabetic rats. Jpn. Pharmacol. Ther. 65, 2888–2894. doi: 10.1021/acs.jafc.6b05627
Tseng, C.-W., Liao, C.-Y., Sun, Y., Peng, C.-C., Tzen, J. T. C., Guo, R.-T., et al. (2014). Immobilization of clostridium cellulolyticum D-psicose 3-epimerase on artificial oil bodies. J. Agric. Food Chem. 62, 6771–6776. doi: 10.1021/jf502022w
Ung, Y. T., Ong, C. E., and Pan, Y. (2018). Current high-throughput approaches of screening modulatory effects of xenobiotics on cytochrome P450 (CYP) enzymes. High Throughput. 7:29. doi: 10.3390/ht7040029
Wagner, N., Bosshart, A., Failmezger, J., Bechtold, M., and Panke, S. (2015). A separation-integrated cascade reaction to overcome thermodynamic limitations in rare-sugar synthesis. Angew. Chem. Int. Ed. Engl. 54, 4182–4186. doi: 10.1002/anie.201411279
Wei, M., Li, Z., Li, T., Wu, B., Liu, Y., Qu, J., et al. (2015). Transforming flask reaction into cell-based synthesis: production of polyhydroxylated molecules via engineered Escherichia coli. ACS Catal. 5, 4060–4065. doi: 10.1021/acscatal.5b00953
Windle, C. L., Müller, M., Nelson, A., and Berry, A. (2014). Engineering aldolases as biocatalysts. Curr. Opin. Chem. Biol. 19, 25–33. doi: 10.1016/j.cbpa.2013.12.010
Wong, J. X., Ogura, K., Chen, S., and Rehm, B. H. A. (2020). Bioengineered polyhydroxyalkanoates as immobilized enzyme scaffolds for industrial applications. Front. Bioeng. Biotechnol. 8:156. doi: 10.3389/fbioe.2020.00156
Wu, Y., Chen, T., Liu, Y., Tian, R., Lv, X., Li, J., et al. (2019). Design of a programmable biosensor-CRISPRi genetic circuits for dynamic and autonomous dual-control of metabolic flux in Bacillus subtilis. Nucleic Acids Res. 48, 996–1009. doi: 10.1093/nar/gkz1123
Wu, S., and Li, Z. (2018). Whole-cell cascade biotransformations for one-pot multistep organic synthesis. ChemCatChem 10, 2164–2178. doi: 10.1002/cctc.201701669
Xia, Y., Cheng, Q., Mu, W., Hu, X., Sun, Z., Qiu, Y., et al. (2021). Research advances of D-allulose: An overview of physiological functions, enzymatic biotransformation technologies, and production processes. Foods 10:2186. doi: 10.3390/foods10092186
Yang, J., Tian, C., Zhang, T., Ren, C., Zhu, Y., Zeng, Y., et al. (2019). Development of food-grade expression system for D-allulose 3-epimerase preparation with tandem isoenzyme genes in Corynebacterium glutamicum and its application in conversion of cane molasses to D-allulose. Biotechnol. Bioeng. 116, 745–756. doi: 10.1002/bit.26909
Yang, J., Zhu, Y., Li, J., Men, Y., Sun, Y., and Ma, Y. (2015). Biosynthesis of rare ketoses through constructing a recombination pathway in an engineered Corynebacterium glutamicum. Biotechnol. Bioeng. 112, 168–180. doi: 10.1002/bit.25345
Yang, J., Zhu, Y., Men, Y., Sun, S., Zeng, Y., Zhang, Y., et al. (2016). Pathway construction in Corynebacterium glutamicum and strain engineering to produce rare sugars from glycerol. J. Agric. Food Chem. 64, 9497–9505. doi: 10.1021/acs.jafc.6b03423
Yoshihara, A., Kozakai, T., Shintani, T., Matsutani, R., Ohtani, K., Iida, T., et al. (2017). Purification and characterization of D-allulose 3-epimerase derived from Arthrobacter globiformis M30, a GRAS microorganism. J. Biosci. Bioeng. 123, 170–176. doi: 10.1016/j.jbiosc.2016.09.004
Zhang, W., Chen, D., Chen, J., Xu, W., Chen, Q., Wu, H., et al. (2021). D-allulose, a versatile rare sugar: recent biotechnological advances and challenges. Crit. Rev. Food Sci. Nutr. doi: 10.1080/10408398.2021.2023091
Zhang, W., Jia, M., Yu, S., Zhang, T., Zhou, L., Jiang, B., et al. (2016a). Improving the thermostability and catalytic efficiency of the D-psicose 3-epimerase from Clostridium bolteae ATCC BAA-613 using site-directed mutagenesis. J. Agric. Food Chem. 64, 3386–3393. doi: 10.1021/acs.jafc.6b01058
Zhang, W., Li, H., Jiang, B., Zhang, T., and Mu, W. (2017). Production of D-allulose from D-glucose by Escherichia coli transformant cells co-expressing D-glucose isomerase and D-psicose 3-epimerase genes. J. Sci. Food Agric. 97, 3420–3426. doi: 10.1002/jsfa.8193
Zhang, W., Li, H., Zhang, T., Jiang, B., Zhou, L., and Mu, W. (2015). Characterization of a D-psicose 3-epimerase from Dorea sp. CAG317 with an acidic pH optimum and a high specific activity. J. Mol. Catal. B Enzym. 120, 68–74. doi: 10.1016/j.molcatb.2015.05.018
Zhang, J., Xu, C., Chen, X., Ruan, X., Zhang, Y., Xu, H., et al. (2020). Engineered Bacillus subtilis harbouring gene of D-tagatose 3-epimerase for the bioconversion of D-fructose into D-psicose through fermentation. Enzym. Microb. Technol. 136:109531. doi: 10.1016/j.enzmictec.2020.109531
Zhang, W., Yu, S., Zhang, T., Jiang, B., and Mu, W. (2016b). Recent advances in D-allulose: physiological functionalities, applications, and biological production. Trends Food Sci. Technol. 54, 127–137. doi: 10.1016/j.tifs.2016.06.004
Zhang, W., Zhang, T., Jiang, B., and Mu, W. (2016c). Biochemical characterization of a D-psicose 3-epimerase from Treponema primitia ZAS-1 and its application on enzymatic production of D-psicose. J. Sci. Food Agric. 96, 49–56. doi: 10.1002/jsfa.7187
Zhu, Z., Gao, D., Li, C., Chen, Y., Zhu, M., Liu, X., et al. (2019a). Redesign of a novel D-allulose 3-epimerase from Staphylococcus aureus for thermostability and efficient biocatalytic production of D-allulose. Microb. Cell Factories 18:59. doi: 10.1186/s12934-019-1107-z
Zhu, Z., Li, C., Liu, X., Gao, D., Wang, X., Tanokura, M., et al. (2019b). Biochemical characterization and biocatalytic application of a novel D-tagatose 3-epimerase from Sinorhizobium sp. RSC Adv. 9, 2919–2927. doi: 10.1039/C8RA10029B
Zhu, Y., Li, Y., Xu, Y., Zhang, J., Ma, L., Qi, Q., et al. (2021). Development of bifunctional biosensors for sensing and dynamic control of glycolysis flux in metabolic engineering. Metab. Eng. 68, 142–151. doi: 10.1016/j.ymben.2021.09.011
Keywords: rare sugars, D-allulose, D-allulose 3-epimerase, biosynthesis, aldolases, applications
Hivatkozás: Chen Z, Gao XD és Li Z (2022) Újabb eredmények a D-allulóz élettani funkcióival és bioszintézisével kapcsolatban. Front. Microbiol . 13:881037. doi: 10.3389/fmicb.2022.881037
Véleményezte:
Youqiang Xu , Pekingi Műszaki és Üzleti Egyetem, Kína;Xinqiang Xie , Guangdong Tudományos Akadémia, Kína.
Copyright © 2022 Chen, Gao és Li. Ez egy nyílt hozzáférésű cikk, amely a Creative Commons Nevezd meg! (CC BY) licenc feltételei szerint kerül terjesztésre . A felhasználás, terjesztés vagy sokszorosítás más fórumokon megengedett, feltéve, hogy az eredeti szerző(k)et és a szerzői jog tulajdonos(oka)t feltüntetik, és hogy az eredeti folyóiratban megjelent publikációt idézik, az elfogadott tudományos gyakorlattal összhangban. Tilos olyan felhasználás, terjesztés vagy sokszorosítás, amely nem felel meg ezeknek a feltételeknek.
*Levelezés: Zijie Li, lizijie@jiangnan.edu.cn
Jogi nyilatkozat: A cikkben kifejtett összes állítás kizárólag a szerzők állítása, és nem feltétlenül tükrözi kapcsolt szervezeteik, a kiadó, a szerkesztők és a bírálók állításait. A cikkben értékelt termékekre vagy a gyártó által tett állításokra a kiadó nem vállal garanciát és nem is támogatja azokat.
